

How Does Heterosis Improve Plant and Animal Breeding?
Plant breeding is the application of genetic principles in developing new plant varieties, known as cultivar development, crop improvement, and seed improvement. Heterosis in plant breeding is described as the superiority of an F1 hybrid over both parents in terms of yield or other characteristics. Heterosis contributes to increased vigour, size, growth rate, yield, or other attributes. However, in exceptional cases, the hybrid may be inferior to the weaker parent. The methods of estimation of heterosis and the genetic basis of heterosis are described here.
Heterosis Definition
Heterosis refers to the superiority of F, hybrids over their parents in one or more characteristics. The word hybrid vigour is a synonym for heterosis. George Harrison Shull coined the term heterosis in 1914.
Some features of heterosis are described below.
Superiority Over Parents: Heterosis results in superiority over its parents in adaptability, yield, quality, disease resistance, maturity, and general vigour. Positive heterosis is often seen as desirable. However, in some circumstances, negative heterosis is preferable. Negative heterosis for plant height, maturity time, and hazardous chemicals, for example, is beneficial in many circumstances since it demonstrates superiority over the parents. In most agricultural plants, heterosis of 40% or more over the superior parent is regarded as substantial from a practical standpoint.
Confined to F1: Heterosis is restricted to the F1, resulting in the production of a cross. As a result of segregation and recombination, it diminishes and vanishes in F1 and later generations of a cross. As a result, heterosis is linked to the F1 generation.
Genetic Control: Nuclear genes regulate the expression of heterosis. In certain cases, heterosis is caused by the interaction of nuclear genes and cytoplasm.
Reproducible: Once recognised, heterosis may be easily reproduced in a specific environment. The expression of heterosis, on the other hand, is more evident in the region of hybrid adaptability.
Relationship with SCA: Heterosis shows a positive relationship with specific combining ability (SCA) variation. The SCA is a measure of dominance variance, and having a high degree of dominance variance is required to carry out a heterosis breeding program.
Heterozygosity Effect: The degree of heterosis is related to heterozygosity since dominance variance is related to heterozygosity. The dominance effects should be most significant in cross-pollinated species and least significant in self-pollinated species. As a result, heterosis occurs more frequently in cross-pollinated crops than in self-pollinated crops.
Masks Recessive Genes: When there is heterosis, the beneficial influence of dominant genes masks harmful recessive genes. As a result, recessive mutant genes are concealed in heterozygous individuals.
Low Frequency: The frequency of good heterotic pairings is quite low. Only a few good heterotic pairings are discovered after screening thousands of F1 crosses. All of the F1 crosses lack desired heterosis.
Genetic Basis of Heterosis
To explain the mechanism of heterosis, two significant theories have been suggested. The first is the dominance theory, while the second is the overdominance hypothesis. Epistasis is also probably related to heterosis. As a result, there are three potential genetic origins of heterosis, which are:
Dominance
Overdominance
Epistasis
Dominance Hypothesis
Davenport (1908), Bruce (1910), and Keeble and Pellew (1910) proposed this hypothesis. This is the most commonly accepted explanation for heterosis. According to this theory, heterosis is caused by the superiority of dominant alleles when recessive alleles are harmful. The hybrid shows heterosis because the deleterious recessive genes of one parent are concealed by the dominant genes of the other parent. Both parents have different dominant genes.
Assume one parent's genetic make-up is AABBccdd and the other's is aabbCCDD. A hybrid of these two parents will have four dominant genes, giving it superiority over both parents having two dominant genes. Thus, heterosis is proportional to the number of dominant genes contributed by each parent.
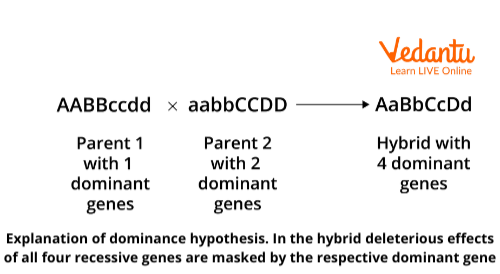
Dominance Hypothesis
Overdominance Hypothesis
Shull and East separately presented this hypothesis in 1908. This hypothesis is known as stimulation of heterozygosis, cumulative action of divergent alleles, single-gene heterosis, super-dominance, and overdominance. Even though Shull and East proposed this hypothesis in 1908, Hull used the word overdominance in 1945 when working on maize. According to this theory, heterosis is caused by the heterozygote's superiority over both of its homozygous parents. Thus, heterosis is proportional to heterozygosity.
The superiority of the heterozygote over both homozygotes may result from:
The production of a superior hybrid substance in the heterozygote that is entirely different from either of the homozygous.
Greater buffering capacity in the heterozygote due to cumulative action of divergent alleles or stimulation of divergent alleles. East explained this theory in 1936, suggesting a set of alleles a1, a2, a3, and a4 with steadily increasing divergence in function. As a result, a combination of more divergent alleles will have more heterosis than a combination of less divergent alleles. Combinations of a1a4, for example, demonstrate more heterosis than combinations of a1a2, a2a3, and a3a4. Overdominance has been reported in barley.
Epistasis
The interaction of alleles from two or more distinct loci is referred to as epistasis. It is sometimes referred to as nonallelic interaction. Non-allelic interactions are classified into three types: additive x additive, dominance x dominance, and additive x dominance. It is widely documented that the presence and size of non-interaction have a positive relationship with the incidence and magnitude of heterosis. Epistasis, especially dominance effects (dominance x dominance), may lead to heterosis. Cotton and maize have both shown this (Moll and Stuber 1974). Various biometrical models can detect or estimate epistasis.
Methods of Estimation of Heterosis
Heterosis is estimated in three ways:
Over mid parent
Over better parent
Over a commercial hybrid
Thus, based on estimation, heterosis is classified into three types, as shown below.
Average Heterosis: When the heterosis is estimated over the mid parent, i.e., the average value of the two parents, it is known as average heterosis, which is calculated as Average Heterosis= {(F1-MP)/MP} X 100
Where F1 is the mean value of F1 and MP is the mean value of the two parents involved in the cross.
Heterobeltiosis: It occurs when the heterosis is estimated to be superior or better than the superior or a better parent. It is known as heterobeltiosis. It is calculated as follows:
Heterobeltiosis= {((F1-BP)/BP) X 100}
BP is the mean value (across replications) of the cross's better parents.
Useful Heterosis: Meredith and Bridge coined the term useful heterosis in 1972. It is also known as economic heterosis and refers to F1's superiority over the normal commercial check type. This sort of heterosis has direct use in plant breeding. It is calculated as follows.
Useful heterosis= {((F1-CC)/CC) X 100}
Where CC is the mean value (across replications) of the local commercial hybrid. Over the conventional commercial hybrid, heterosis is sometimes worked out.
Standard Heterosis: Heterosis is estimated in crops where hybrids are already available for comparison. Standard heterosis refers to this sort of heterosis. This is also directly applicable in plant breeding. It is calculated as follows.
{(F1-SH)/SH} X 100 = Standard heterosis
Where SH is the mean value of the standard hybrid.
Conclusion
Heterosis refers to the superiority of F1 hybrids in one or more characteristics over their parents. The term hybrid vigour is used interchangeably with heterosis. Dr. G. H. Shull coined the term "heterosis" in 1914. Heterosis is the process by which a less vigorous organism is turned into a more vigorous organism by absorbing DNA from the media.


FAQs on Heterosis: Definition, Genetic Basis, and Methods
1. What exactly is heterosis in biology?
Heterosis, more commonly known as hybrid vigour, is a genetic phenomenon where the offspring of a cross between two genetically distinct parents (the F1 hybrid) shows qualities superior to both of its parents. These superior traits can include increased size, growth rate, fertility, yield, or resistance to diseases.
2. What are some common examples of heterosis used in agriculture?
Heterosis is widely used in agriculture to boost crop productivity. A classic example is hybrid corn (maize), which produces significantly higher yields than its parent inbred lines. Other examples include:
- Increased fruit size and yield in tomatoes.
- Enhanced disease resistance and yield in rice varieties.
- Superior fibre quality and yield in hybrid cotton.
3. What is the primary difference between heterosis and inbreeding depression?
Heterosis and inbreeding depression are essentially opposite phenomena. Heterosis results from an increase in heterozygosity (having different alleles for a trait) when two diverse parents are crossed, leading to enhanced traits. In contrast, inbreeding depression is the result of an increase in homozygosity (having identical alleles) from mating closely related individuals, which often leads to a reduction in fitness and the expression of harmful recessive traits.
4. How do scientists explain the genetic basis of heterosis?
There are two main hypotheses that explain why heterosis occurs:
- The Dominance Hypothesis: This theory suggests that each parent carries different harmful recessive alleles. In the hybrid offspring, the dominant, beneficial alleles from one parent mask the harmful recessive alleles from the other parent, resulting in overall superior fitness.
- The Overdominance Hypothesis: This theory proposes that the heterozygous state (e.g., Aa) is intrinsically superior to either homozygous state (AA or aa). The combination of two different alleles at a single locus produces a synergistic effect that enhances the trait beyond either parent's contribution.
5. Why does the effect of hybrid vigour often disappear after the F1 generation?
The superiority of heterosis is most pronounced in the first filial (F1) generation because this generation is uniformly heterozygous for the key traits. When F1 individuals are crossed or self-pollinated to produce an F2 generation, Mendelian segregation and recombination break up these favourable heterozygous gene combinations. This leads to an increase in homozygosity, causing the superior traits to diminish and the population to become less uniform.
6. Is heterosis always a positive phenomenon in plant breeding?
While heterosis is usually sought for positive traits like higher yield (positive heterosis), it can also manifest as negative heterosis, which can be desirable in certain situations. For example, a breeder might want negative heterosis for:
- Plant Height: Shorter, sturdier plants are less prone to wind damage.
- Maturity Time: Earlier maturity can be advantageous for fitting into specific cropping seasons.
- Toxic Compounds: Reduced levels of naturally occurring toxic substances (like glucosinolates in rapeseed) are beneficial.
7. How is heterosis measured in practical breeding programs?
Heterosis is typically estimated in a few key ways to determine its practical value:
- Average Heterosis (Mid-parent): The performance of the F1 hybrid is compared to the average performance of its two parents.
- Heterobeltiosis (Better-parent): The F1 hybrid's performance is compared specifically to the parent with the superior performance for that trait.
- Economic or Useful Heterosis: The F1 hybrid is compared against the current best commercial variety available in the market. This is the most important measure for practical application and commercial success.
8. How is the concept of heterosis applied differently in animal breeding compared to plant breeding?
While the underlying genetic principle is the same, the application differs. In plant breeding, heterosis often focuses on traits like yield and disease resistance. In animal breeding, heterosis is also evaluated for more complex traits. For instance, maternal heterosis refers to the improvement in a crossbred mother's reproductive and maternal abilities (like milk production and fertility), while paternal heterosis can refer to improved reproductive traits in crossbred males, such as fertility and libido.





